
Големи структури на
ЦНС – структура и функция
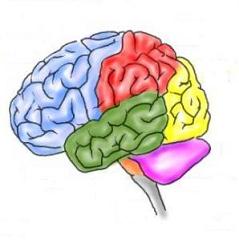
Главният мозък е
разположен в черепната кухина и външната
му форма съответства на сложните й
очертания. Частите на главния мозък са
се развивали последователно и свързано
с усложняване на функциите му и са:
1. Продълговат мозък –
Medulla oblongata
2. Заден мозък –
Metencephalon с две части: мост
– pons и малък мозък –
cerebellum
3. Среден мозък –
Messencephalon
4. Междинен мозък –
Diencephalon (Thalamus
и Hipothalamus)
5. Краен мозък -
Тelencephalon (Hemispheria
Cerebralis и Corpuscallosum)
6. Гръбначен мозък –
Medulla Spinalis
7. Подкорови сиви ядра.
Medulla
oblongata – първата и
най-ниско разположена част на главния
мозък и е непосредствено продължение
на гръбначния мозък. Продълговатият
мозък е една от частите на ЦНС, чиято
функция е свързана с регулацията на
основните жизнени процеси – дишане,
съдов тонус, контрол в/у сърдечната
дейност и др. Тук се намират ядрата
(струпвания на сиво вещество) на
черепномозъчни нерви, осигуряващи
интервенцията на вътрешни органи и
органи на кръвообращението. Чрез него
се осъществява връзката между всички
по-висшестоящи части на нервната система.
На повърхността на продълговатия мозък
се появяват 4 от гръбначно-мозъчните
нерви – n.glosspharyngeus
(glossos - език, phrungs
- глътка), часто то n.accessorios,
n.vagus
(блуждаещ) – инервира сърцето, а в
браздата пред олива (маслина) се намират
коренчетата на n.hypoglossos
(подезичен) и ретикуларна формация. При
прекъсване на tractus
corticobularis се получават
парализи на езика.
PONS –
мостът е долната част на задния мозък
и се разполага между продълговатия и
средния мозък. Два пъти по-голям е от
предния мозък. В моста се съдържат освен
бяло вещество и ядра от сиво мозъчно
вещество – nuclei pontis
(ядра на моста). Всички аферентни и
еферентни пътища минават през него, но
са пръснати и част от тях се отклоняват
в различни пътища. Тези пътища носят
информация от моста към малкия мозък,
както и такива, които носят информация
от кората към подкорието. От моста водят
началото си n.trigeminus
(троичния нерв – осигурява общата
сетивна инервация на по-голямата част
от главата и двигателната инервация на
дъвкателните мускули), n.facialis
(лицев нерв – инервира мимическата
мускулатура на лицето), n.abdeucens
и n.statoacusticus
(състои се от две части: слухова и
равновесна. Първата е свързана с
приемането и провеждането на слуховите
усещания от охлюва на вътрешното ухо,
а втората провежда нервните импулси от
преддверието на вътрешното ухо, които
са свързани с контрола, регулацията на
положението и движенията на тялото.)
Патологични изменения – нарушена
трофика – хранене, както и прекъсвания
на пътищата и нервите от различен
произход водят до промени в инервираните
от съответните нервни зони. Напр. при
засягане на n.facialis
след излизнето му от черепната кухина
се наблюдава едностранна паралиа на
лицевата мускулатура, ако патологията
е по пътя от ядрата до мястото, където
нервът напуска понса, се наблюдава
частична възможност за движенеи на
лицевите мускули, поради прекръстосване
на влакна от лявата и дясната половина
– болният може да сбърчи челото. Това
е диференциално-диагностичен признак
между централна и периферна парализа
на n.facialis.
Трите нерва – n.facialis,
n.trigeminus и
n.statoacusticus
напускат понса през nomo-церебралния
ъгъл. Данни за увреда на трите нерва
едновременно е патогномотичен (определящ)
признак за Neurinoma – туморен
процес, чиято локализация е извън мозъка
и ранното му диагностициране е от
изключителна важност. Един от трите
очедвигателни нерва се намира в понса.
И тук има ретикуларна формация.
MESSENCEPHALON
– средният мозък е по-малка структура
с по-малко количество нервни пътища
(аферентни и еферентни). В началото се
развива във връзка с органа на зрението,
но по-късно във филогенетичното развитие
на предния мозък и други части на ЦНС в
него се появяват и нови структури –
струпвания от сиво мозъчно вещество и
снопове от проводни пътища към
по-високостоящите части на мозъка –
междинния мозък и голямо-мозъчните
хемисфери. Дорзапната му (гръбначна)
повърхност има своеобразни очертания.
На нея се намират 4 възвишения – colliculi
(хълмче), superior (горно) и
inferior (долно), по едно горно
и долно за всяка половина. Горните са
ангажирани със зрителната информация,
а долните със слуховата. От средния
мозък излизат n.ocomotorius
и n.trohlearis,
които са свързани с волевите движения
на очните ябълки. При патологични промени
(от кома до сънливост, т.е. промени
свързани със съзнанието) се наблюдава
блокиране на движенията на очните
ябълки. И тук има ретикуларна формация.
CEREBELLUM
– малкият мозък е разположен в задната
черепна ямка. Неговата функция е свързана
основно с равновесието и координацията
на движенията на тялото. При човека, при
когото двигателната функция е силно
развита и малкият мозък е най-силно
развит. Той е съставен от една средна
част, която според своя вид носи названието
червей (vermis) и две силно
развити странични части: полукълба на
малкия мозък – hemispheria
cerebelli – свързан е със
съседните части на ствола чрез снопове
от влакна, наречени крачета – pedunculi
cerebellaris. Нагоре той се
свързва със средния мозък чрез горното
краче – pedunculus cerebellaris
superior, а с продълговатия
мозък чрез долното краче - pedunculus
cerebellaris inferior.
Ядрата от сивото мозъчно вещество, по
4 от всяка страна, са разположени сред
бялото вещество, те са: nucleus
detantus (зъбчато ядро), nucleus
emboliformis (капковидно),
nucleus globosus
и nucleus fastigii.
Кората на малкия мозък
се характеризира с това, че във всички
свои части има еднообразна структура,
свързана вероятно с еднообразната
функция. На нея се различават 3 слоя:
един повърхностен, наречен така поради
микроскопския си вид – молекулярен,
вътрешен – зърнист, и среден слой на
клетките на Purkinje. Тези
клетки са с голямо крушовидно тяло.
Техните дендрити достигат до повърхностния
слой, а от вътрешната страна на клетките
излизат миелинови влакна, които се
насочват към вътрешността в бялото
мозъчно вещество. Веднага след началото
си нервните клетки на Purkinje
дават разклонения, което осигурява
включване във функция на множество
клетки. Малкият мозък на човека на база
на филогенетичното му развитие се
разделя на най-стара част – archicerebellum,
стара част paleocerebellum и нова
част neocerebellum. При увреждане
на най-старата част, най-често при децата
настъпва разстройство на равновесието.
Болният не може да поддържа равновесие
в изправено положение, докато в легнало
състояние не се откриват съществени
смущения в координацията на движенията.
По отношение на старата част при човека
се знае по-малко. От опити при животни
се знае, че тази част е свързана с
поддържане на определено положение на
тялото и с движение на крайниците. Новата
част на кората на малкия мозък е под
контрола на голямомозъчните хемисфери.
Тя играе основна роля при координирането
на диженията особено на крайниците и
осигурява синергизма на мускулното
действие. Движенията са разстроени и
несигурни. Тази част има отношение и
към мускулния тонус. При нарушение се
появява разстройство в синергичното и
последователно функциониране на
мускулите, както и намаляване на мускулния
тонус – от хипотония (намален мускулен
тонус) до пълна атония (липса на такъв).
Ganilion
Basalis – тази структура е
представена от putamen,
palidum, substantia
nigra, stiamum.
Всички участват в контрола на двигателните
процеси. При нарушение на субстанция
нигра и стриатима се развина синдрома
на Паркинсон. Има намалено количество
допамин. На лице са следните симптоми
– треперене, мускулна регидност, поради
повишен мускулен тонус. Симптом на
зъбчатото колело, брадилалия, брадипсихия
и др. Тези симптоми могат да се наблюдават
и при съдови увреждания.
DIENCEPHALON
– разположен е между базалните части
на краймозъчните полукълба. Състои се
от две главни части: таламов мозък и
подхълмие. Таламовият мозък се състои
от 3 части: thalamus, metathalamus,
epithalamus. Вътрешното
устройство на таламуса е сложно, има
сложни многообразни функции. Играе роля
на междинна станция, в която нервните
импулси идват от определени мозъчни
структури, претърпяват междинна обработка
и се препращат към други. В 3 отделни
части на таламуса се описват над 20 ядра,
свързани чрез сложни връзки помежду си
и с различни части на мозъка. Ядрата в
предната му част са свързани с обонянието,
ядрата в междинната му част имат сложни
връзки с други части на самия таламус
и с части на кората на хемисферите.
Метаталамуса има отношение към предаване
на информацията, свързана със слуха и
зрението. В епиталамуса има зона свързана
с обонянието. Друга част на междинния
мозък е хипоталамусът. Тук в две телца
(corpora mamillaria)
се намират подкоровите центрове на
обонятелния мозък. Chiasma
opticum е място, където част
от влакната на зрителния нерв се
прекръстосват с тези от противоположната
страна и се отправят към противоположната
страна на мозъка. Хипофизата е една от
жлезите с вътрешна секреция. Синтезира
се растежният хормон. Основните функции
на хипоталамуса са свързани с регулацията
на телесната температура, отделянето
на вода от бъбреците и др.
Част от мозъчните
структури са и мозъчните стомахчета,
които са пълни с гръбначно-мозъчна
течност (ликвор) и играят роля на
своеобразен буфер при някой възпалителни
заболявания и травми върху мозъка. По
състоянието на ликвора се съди за
определена патология (гноен менингит).
Кръвоснабдяването е от особена важност
за правилното функциониране на мозъка.
То се осъществява от две чифтни артерии
– a.vertebralis
за тилната област и a.carotis
komunis. Лявата и дясната
a.vertebralis се
сливат в една нечифтна a.basiralis.
Преди сливането се отделят къси клончета
за кръвоснабдяване на продълговатия
мозък и др. за кръвоснабдяване на малкия
мозък. От вътрешната сънна артерия се
отделят клончета, които кръвоснабдяват
средния и преден дял на мозъка.
TELENCEPHALON
– краен мозък, съставен от две
голямо-мозъчни хемисфери – hemispheria
cerebriл свързани помежду
си чрез мазолестото тяло (corpus
callosum). Крайният мозък се
развива във връзка с обонянието – това
е неговото място и значение при най-нисшите
гръбначни животни. Постепенно към него
се изместват най-висшите функции на
нервната система, което определя
поведението на животното. В сивото
вещество на хората на хемисферите се
извършва най-висш анализ и синтез на
информация, идваща от аференти пътища.
Тук е локализирана условно рефлексната
дейност на нервната система. Чрез нея
се определя поведението на индивида.
Повърхността на голямомозъчните
хемисфери е неравна. На нея се намират
множество различно дълбоки бразди –
sulci cerebri,
които отделят от повърхността извивки
с различна форма и посока, наречени
gyris cerebri.
По-основните от тях са: sulcus
centralis, s.precentralis,
s.postcentralis,
с едноименните им гируси (гънки), sulcus
infraparietalis, fissure
longitodinalis cerebri,
разположена между двете хемисфери,
страничните фисури (бразди) – temporalis
(слепоочна) superior, temporalis
medius, temporalis
inferior и temporalis
transverza, отделяща предния
от малкия мозък. Чрез нагъването
повърхността, а от там големината на
кората и сивото мозъчно вещество се
увеличава многократно.
CORTEX
– сиво вещество, мозъчна кора. Сивото
вещество, където и да се намира, за него
е приоритет, че има нервни клетки с тела,
а бялото вещество се състои от различни
израстъци на нервни клетки. В неокортекса
нервните клетки се разпределят в 6
пласта: 1.полиморфен слой – повече глия;
2.дребни зърнести клетки; 3.дребни
пирамидни клетки (2 и 3 са с множество
дендрити и къси аксони); 4.едрозънести
клетки – с множество дендрити и по-къси
аксони; 5.големи пирамидни клетки на
Бетс (митрални) – клетки с най-дълги
аксони и дендритни разклонения; 6.вътрешен
слой – състои се от разнообразни неврони,
малко на брой – кандиларни, звездовидни
– полиморфни, както за първия, така и
за шестия слой има малко информация. В
палеокортекса има три слоя: 1.полиморфен
слой – повече глия; 2.различни неврони
със зърнеста и пирамидална структура;
3.състои се от разнообразни неврони,
както шести слой на неокортекса. Разликата
между палеокортекса и архикортекса е,
че при архикортекса има два слоя.
Най-диференцираната
част на мозъчната кора е кората на
предните мозъчни полукълба. ЦНС е покрита
с мозъчни обвивки – менинги. Те са 3
вида:
А.Duramatter
– най-външна, прилепнала към черепа,
като го тапицира отвътре с единия си
пласт. Между двата пласта има много
кръвоносни съдове. Под duramatter
има субдурално пространство. Играе
ролята на буфер при удар. При възрастни
хора при лек удар може да се получи
субдурален хематом.
Б.Arachnoidea
– паяжиновидна мрежа – има структури,
които произвеждат ликвор. Под arachnoidea
има субархинодипно пространство.
Най-честата причина за епилепсия е
нарушението на това пространство.
В.Pia matter
– мека мозъчна обвивка – прилепнала
плътно към най-външния слой на неокортекса
и следва гънките на мозъчната кора.
Подкорови сиви ядра
– ганглии (базални ганглии) – сред
бялото вещество на telencephalon-а
има струпвания от сиво вещество. Базалните
ганглии са различни по форма и с различни
функции. Свързани са с двигателните
процеси. Най-голямото ядро е Putamen,
следват го Palidum Globus,
Substantia Nigra
– черно ядро (ако има увреда – силно
намаляване на допамин – следва
Паркинсонизъм). Смущават се движенията
поради тремор с регидно повишен мускулен
тонус. Синдромът се описва като броене
на монети. Има наследствена паркинсонова
болест. Българинът Иван Раев открива,
че Паркинсон се лекува с Белла Дона.
Болестта може да се появи и при
артериосклероза. От там идва понятието
брадикинезия (забавеност на движенията,
хипотомия – липсваща мимика на лицето
и др.).
Nucleus
venticularis – ядро, участващо
в контрола на двигателните процеси.
Свързва се с черната жълтеница у децата
(след раждането). Идва от RH-несъвместимост
у родителите. Получава се много тежък
синдром на нарушение на движенията –
хиперкинези – не могат да ходят, да
вършат нищо. Хепатолентикуларен синдром
– дължи се на смущение на обмяната на
мед в организма. Смущение на движенията
у млади хора (хиперкинези). Тази болест
е необратима. Horea minor
– появява се в детска възраст – непослушно
дете, недисциплинирано. Дължи се на
ревматичен процес, засягащ подкоровите
ядра. Ако се открие навреме, се лекува.
Жънтингтонова болест – наследствена,
като няколко члена на 1 фамилия след
40-50 годишна възраст получават паразитни
движения.
MEDULLA SPINALIS
– гръбначен мозък. Дълга напречна
структура в костен канал, с напречно
сечение около 1 см. и се помещава в един
костен канал, образуван от прешлените
на гръбначния стълб. Бялото вещество
се намира отвън на гръбначния мозък, а
сивото вещество е вътре. При напречно
сечение има форма на пеперуда – 2 предни
и 2 задни рога, свързани помежду си. В
центъра има централен канал с мозъчна
течност. След като свърши гръбначния
мозък, в последните структури има
пространство с нервни окончания и
ликвор. Конска опашка. Там се прави
пункция. Функцията на гръбначния мозък
се изучава. Състои се от 31 сегмента.
30-те от тях имат приблизително еднаква
картина на сиво и бяло вещество, а 31-ят
е различен. Функционалната стойност на
тези сегменти е: 8 шийни сегмента, 12
гръдни сегмента – торакални, 5 кръстни
– ломбални, 5 свещени – сакрални. Има
две зони в кръстната област и шията, в
които има повече нервни клетки
(удебелявания). Тези клетки се свързват
с крайниците. Отстрани на гръбначния
мозък има гръбначно-мозъчни възли. Всеки
ганглиум спинали има 1 суниполарен
неврон, има израстък, довеждащ до него
информация от рецептора по крайниците,
кожата и т.н. Има кожни зони, отговарящи
за всяка една от 30-те двойки – дерматоми.
Миотоми – групи мускулни влакна.
Сегментен рефлекс – чрез рефлексна
дъга се достига до един от ломбалните
рефлекси. Спинобулбарните пътища (пътища
за дълбока сетивност) са до продълговатия
мозък. Предават аферентна информация
от ставите. Път за болка и усет (тратус
синусталаминус) – униполарен неврон,
който изпраща своя сом в противополжната
половина на гръбначния стълб до таламуса.
Формиране на пътища за допирна сетивност
(спиноталамичен път). Той се кръстосва
на нивото на всеки сегмент. Сетивността
върви по два пътя – спиноталамичен и
спинобулбарен.
Lemiscus
medialis – кръстовище
на нивото на булбуса. Всяка половина от
гръбначния мозък отнася информация до
главния мозък. Открити са стар и нов
спиноталамичен път (представя информация
за фините движения като писане например),
то се контролира от аферентен път.
Еферентни пътища в
бялото вещество – движат се от кората
към гръбначния мозък – пирамиден път
(големите пирамидални клетки на Бетс
слизат надолу към гръбначния мозък).
Decussatio
pyramida – набраздени
са 2 големи пирамиди на границата между
продълговатия и гръбначния мозък. Има
и некръстосани пирамидни пътища.
Еферентни и аферентни пътища в бялото
вещество. Има един път – rubio
spinale (има ядро), който
изпраща еферентни пътища към гръбначния
стълб, поддържа равнището на мускулния
тонус.
Reticullo
spinalis – Основни са
пътищата за сетивността и за двигателните
неврони.
В мотоневроните в
предните рога на гръбначния мозък може
да има засягане на самите аксони и се
получава полиомиелит. Периферен
двигателен неврон – засегната е самата
клетка в предните рога. Характерно е,
че изчезва съответният сегментен
рефлекс, мускулният тонус отслабва,
настъпва атрофия на мускулатурата.
Мотоневроните приемат команди от
мозъчната кора. Всичко се контролира
от неокортекса. Ако този контрол е
нарушен, сегментният рефлекс става
болезнен или особено усилен. Централна
парализа – увреда на централен двигателен
неврон. Появяват се патологични,
филогенетични стари рефлекси – рефлекс
на Бабински – например ако се дразни
кожата на ходилата, се получава повдигане
на палеца нагоре. Нарушена е миелинизацията
на пътищата. Спастично повишен мускулен
тонус – стегната мускулатура – синдром
на затварящото се джобно ножче.
Ретикуларна
формация и лимбична система
В централната част на
шийния сегмент на гръбначния мозък
съществуват клетки, които са разпръснати
или групирани в малки ядра с различна
големина. Характерно за тези клетки е
наличието на слабо разклонена мрежа от
дендрити и наличието на двуполюсни
аксони – с възходящ и низходящ клон,
които дават разклонения и осъществяват
връзка с множество други неврони. Това
натрупване на клетки с гъсто преплетени
нервни влакна, имащи ход с различно
направление, образува не толкова
структурно, колкото функционално
обединение, което е познато като
ретикуларна формация (РФ). Тя се
разполага по цялото протежение на
мозъчния ствол, като прониква и в
гръбначния мозък. Тя се състои от клетки
с различна големина и форма, които са
разпръснати по една или групирани в
множество ядра. Техните дендрити се
разклоняват много слабо, а аксоните им
много силно. Последните се делят на
възходящ и низходящ клон, от който
излизат многобройни колитерали,
осъществяващи контакт с огромен брой
неврони. Нервните влакна вървят в
най-различни направления и осъществяват
разнообразни връзки. РФ е свързана с
всички отдели на ЦНС. Нейните влакна
са: ретикулопетални, ретикулофугални
и ретикулоретикуларни. Към ретикулопеталните
влакна се отнасят такива, идващи от
аферентни пътища на мозъчния ствол, от
малкия мозък, от вегетативните пътища
на мозъчния ствол, от малкия мозък, от
вегетативните подкорови центрове и от
цялата краймозъчна кора. Чрез тях
ретикуларните клетки получават импулси
от почти всички части на ЦНС.
Ретикуларнофугалните влакна се насочват
от клетки на ретикуларната формация
към краймозъчната кора, ядрата на
черепномозъчните нерви, предните колонки
на гръбначния мозък и главния мозък.
Ретикулоретикуларните влакна свързват
отделни ядра и отделни клетки на РФ.
Открива се наличието на възходяща
подтискаща система. Поддържа тонуса на
мозъчната кора. На това се дължи
бодърстването (араузъл). Тя представлява
сложен рефлексен център. Счита се, че
ядрата на РФ вземат участие във всички
сложни рефлекси, изискващи участието
на много мускули в разнообразно съчетание
(говор, гълтане, дишане и т.н.). При
патология на низходящата активираща
система се наблюдава нарколепсис
(непрекъснато заспиване, енопсия –
липса на сън). Недоспиването говори за
недобре оформена РФ.
Лимбична система
(ЛС) се използва за означаване на
най-старата във филогенетично отношение
част на мозъчната кора. Анатомично тя
включва: gyrus paraterminalis
(g.subcalozum),
g.cinguli,
g.hyppocampi,
nucleus amygdalae.
През 1930-33 Папец открива и двустранна
взаимовръзка между таламус и хипоталамус
(hyppocampus, nucleus
amygdalae, corpus
mamillariae, fornix).
По такъв начин се формира затворена
система, която е известна като кръг на
Папец.
Hyppocampus е
склад на паметта, най-напред тя се
складира там. Nucleus amygdalae
отговаря за агресивното поведение
(оперативно се премахва). До неотдавна
се е поддържало схващането, че функциите
на ЛС са свързани с вегетативните функции
– повлияване на кръвно налягане, дишане,
моториката на храносмилателната система,
зенични реакции. Днес е ясно, че ЛС не е
вегетативен мозък. Корсаков открива,
че при алкохолици corpus
mamillariae са увредени. Има
амнезия. При хипокампотомия, при лечение
на епилепсия, също има паметови нарушения.
Прави извод, че ЛС участва в организирането
на паметта, а тя се организира и от всички
части на мозъка. Gyrus cinguli
е дълга, голяма гънка на мозъчната кора,
над мазолестото тяло. Свързва 4-те дяла
на мозъка, тя непосредствено е свързана
с ЛС. Цялата челна част на неокортекса
контролира ЛС. Gyrus
parahyppocampi – пак контролира.
Функционални
зони на мозъчната кора
Кората на церебралните
хемисфери е най-младата и най-съвършената
структура на нервната система. Тук са
зоните, които са сензорни входове и
оторни изходи на кората, и се обозначават
като първични. Първоначално е направен
опит мозъчната кора да се раздели на
цифрови полета, но днес използваме
картата на Бродман. Тези полета са
коригирани (цифрови полета). Наред с тях
се прави и друго разпределение на
функционалните зони (фз), които могат
да включват повече зони – първични,
вторични и третични. За част от тях
функциите са доказани. В тях съответната
функция по правило е соматотопично
представена точка по точка, запазвайки
пространственото разположение на
периферните рецептори и мускули.
Уврежданията на първичните полета водят
до просто блокиране на входовете и
изходите на точно ограничени анестезии,
слепи участъци, парези и др. Следователно,
това са корови зони, които изпълняват
елементрани, а не висши функции и те са
генетично кодирани за своята дейност.
В непосредственото обкръжение на
първичните полета се намират корови
зони, които запазват определена свързаност
с нейната сетивна или двигателната
функция, но извършват по-сложни операции
за преработка на сензорната информация
(гнозис или перцепция) и за програмиране
на двигателни действия (праксис). Това
са вторични корови полета, чието увреждане
не води до анестезии и парализи, а до
нарушаване на познавателната (когнитивната)
сетивната преработка (агнозии) и до
разрушаване на изградени двигателни
навици и сръчности (апраксии).
В неокортекса има и
зони, които не са свързани с никакъв вид
определена сетивна преработка, нито с
двигателни команди. Тук спадат „зоната
на трите лоба”, т.е. граничната област
на теменния, тилния и слепоочния дял
(най-вече 37о поле по Бродман), както
т.нар. префронтална кора – челната кора,
разположена напред от премоторната
зона.
Първични ФЗ (ПФЗ):
ЗРИТЕЛНА ПФЗ – тя
е модалностно специфична – 17о поле по
Бродман. Кората е със силно развит
четвърти слой едрозърнести клетки,
разпределени по определен ред, наречен
ретинотопия. Тази зона е около fisura
calcarina. В левия и десния
дял тази зона има симетрия. Хемианопсия
– централна повреда в стриалната кора,
може да се вижда само в ляво и дясно.
СЛУХОВА ПФЗ – 41о
поле по Бродман. Тя е модалностно
специфична. Намира се до sulcus
silvii. Невроните са повече
от четвърти слой. Има тонотопия: медиално
за високи, латерално за ниски честоти.
Наблюдава се симетрично функциониране.
СОМАТОСЕНЗОРНА ПФЗ
– модалностно специфична за сетивността
на цялото тяло. Обхваща почти изцяло
задната централна гънка. 3-о поле по
Бродман. Точно разпределение на невроните
от Пенфилд (човече на Пенфилд – доказва
соматотопия – разфасовано човече).
Соматотопията на тези клетки е посочена
от Джексън още през 19 век – описал е
местни сетивни Джексънови епилептични
пристъпи, които могат да започват от
устата, лицето и т.н. Симетрично
функциониране, има т.нар. хемианестезия
и др.
МОТОРНА ПФЗ – намира
се в gyrus precentralis,
4о поле по Бродман. Тя е еферентна с
развит пети слой клетки – големи
пирамидни клетки на Бетс. Чрез тях се
изпращат двигателни команди, в нея тези
клетки са разпределени соматотопично
и освен това има симетрично функциониране.
Тук се открива начало на кръстосани
кортикобулбарни свойства. Хемипарализа
– при нарушение в срещуположната гънка,
ако и тя се припокрива по симптоми –
знае се от патологията.
Вторични асоциативни
зони (ВАЗ):
ЗРИТЕЛНА ВАЗ –
включва 18о и 19о поле по Бродман и се
намира в lobus occipitalis.
Тя е модалностно специфична. Втори и
трети пласт клетки са интензивно развити,
няма доказателство за ретинопатия.
Получава информация от подкоровите
структури. Има симетрично и относително
асиметрично функциониране. Буквите
като основен езиков стимул се разпознават
по-добре от тази зона на зрителното
полукълбо. При нарушение се получава
буквена агнозия – доказателство за
асиметрична функция на ЗВАЗ. Има
зрително-модалностна обработка.
СЛУХОВА ВАЗ – намира
се в слепоочния дял, включва 42, 22 и 21 поле
по Бродман, по-сложна е функцията на
невроните. Анатомично, тази зона се
характеризира с това, че вторият слой
е най-силно развит. Информацията постъпва
от подкорови зони. Няма тонотопия. При
увреждане се наблюдава тихотония,
фонемна агнозия – нарзпознаване на
звуците. Има асиметрия (известна).
СЕНЗОРНА ВАЗ –
представена е в 1 и 2 поле и задна париетална
кора – 5 и 7 поле по Бродман. Микроскопски
има развити трети и четвърти слой клетки.
Получава информация от подкорови
структури (таламус). Наблюдава се
значителна асиметрия. По принцип тази
зона няма отношение към езика, но чрез
опипване се предава в тактилна модалност
(писаните по кожата букви се разпознават).
Преписва й се сложната сетивност за
вибрации, дискриминационният усет –
изследва се чрез пергел с две остриета,
двумернопространственият усет.
ДВИГАТЕЛНА ВАЗ –
в 6 и 8 поле по Бродман. Намира се в челния
дял. Интензивно развит е пети слой с
пирамидни клетки на Бетс, с къси аксони.
Подкоровите струткури, до които достигат,
са сивите ядра на подкорието. Свързани
са с волевите действия (праксис), за
които човек се обучава в индивидуалния
живот. Имат симетрично функциониране.
Изключение е писането. При нарушение
се получава аграфия. Принципно нарушенията
се наричат апраксии.
Третични ФЗ:
По две във всяко мозъчно
полукълбо – предна и задна третична
зона.
Задна третична зона –
свързана с аферентна информация. Намира
се в края на sulcus silvii
в зона на припокритие. Няма модалност.
Тук се слючва гнозиса, независимо откъде
идва информацията. Има абсолютна
асиметричност. Зрителна, слухова, а от
там и езикова в лява, а в дясна хемисфера
всякаква друга информация. 39, 40 и 42 поле
по Бродман (зона на Вернике) и края на
22 поле са свързани с четенето и писането,
защото има маркиране чрез алексия и
аграфия.
Предните третични зони
са в предните участъци на lobus
frontalis. Най-интензивно
изградена, представена е от 44 поле по
Бродман (зона на Брока), около него е 45
поле по Бродман, свързано с експресията
на езиковата зона. Занимава се с човешката
дейност (писане, говорене, пеене, мислене,
емоции). Това е критична зона.
Функционални
блокове на ЦНС
Вертикалната организация
на нервната система е изградена от три
блока:
Първи блок –
ретикуларна формация (араузъл), включващ
сън и бодърстване. Сънят е жизнена
необходимост за организма, подобно на
храната и кислорода. Сънят е ритмично
настъпваща промяна в активността на
мозъка, съпроводена от относителна
безсънливост. Сънят при човека е
синтезиран с фазите тъмно/светло. Сънят
се съпровожда от рязко намаляване на
двигателната активност, позата при
възможност е легнало положение, очите
са затворени. Първоначалната представа
за съня е свързана с натрупване в мозъка
на фактора на умората, т.е. сънят
представлява почивка за нервната
система. Регулирането на състоянието
на сън и на бодърстване зависят до голяма
степен от взаимодействията между кората
на крайния мозък и структурите в мозъчния
ствол. Два медиатора имат клочова роля
за настъпването на парадоксалния сън
– по време на RGM фазата
се повишава активността на ацетилхолинергични
неврони и се повишава нивото на серотонина.
По време на класическия сън обмяната
на веществата в мозъка е от тип
възстановителна със синтезиране на
енергийни източници. По време на
парадоксалния сън на синаптично ниво
настъпва прехвърляне от краткосрочната
към дългосрочната памет.
Втори блок –
приемане, преработка и съхраняване на
информацията. Приемане чрез сетивата,
преработката се осъществява в мозъчните
подкорови структури, слуховата, зрителната
информация се съхраняват в паметта –
задържането на влязлата информация в
мозъка. Тук се осъществяват взаимовръзки
между различни мозъчни структури и
мозъчната кора. Участва хипокампусът.
При увреда в префронталната зона се
достига до тежка амнезия.
Трети блок –
осъществява организирането и контрола
на движенията. Включва предните зони
от мозъчните полукълба, челния дял и
малка част от задния дял. Първичните
моторни зони, вторичните полета. Праксис
– съвкупност от движения на волеви
действия. Организация от предните
третични зони. До първичните зони
информацията пристига от периферията
или се изпраща. Вторичните връзки се
осъществяват по схемата кора-подкорие.
Третичните връзки са на ниво кора-кора,
асоциативни участъци от неокортекса
нямат връзка с подкоровите неврони от
едната част към другата кора на мозъчното
полукълбо към другата кора на мозъчното
полукълбо. Когато има нарушения на
мозъка в челните дялове може да се получи
разпад на дейността. При увред в лявата
или дясната зона тези нарушения са
различни. Разликата между лявата и
дясната зона е в това, че в дясната зона
няма аспонтанност, а синдром на Мори –
глуповато, безкритично отношение, говори
много, резоньорство.
Перцепция
и агнозии
Гнозисът (перцепцията,
възприятието) може да се дефинира като
възприемане на позната информация
(слухова, зрителна, проприоцептивна,
тактилна, кинестезична) или на нова
информация с възможност да се разпознават
или категоризират отделни признаци или
цели обекти, факти, събития.
Агнозията пък се
разглежда като възможност да се
разпознават сензорни стимули от определен
клас. Пациентът с агнозия има затруднения
при изпълнение на тестове за узнаване,
като в същото време няма сензорни,
вербални и/или интелектуални дефицити.
При разпознаването на обектите могат
да се определят две равнища: разпознаване
на структурата на видения, чутия, опипания
обект от структурите на други вече
познати обекти. Сетивните системи са:
зразителна, слухова, соматосензорна
или тактилна, обонятелна и вкусова.
Сетивните системи са организирани и
описани чрез следния традиционен модел
на Мерцених Каас – 1980 г. Основните пътища
от рецепторните органи стигат (отиват)
към таламус и ядрата в мозъчния ствол,
а от там към неокортекс – първичната
сензорна кора на церебралните хемисфери.
За зрението – окципиталния лоб (17 поле
по Бродман); за слуха – в темпоралния
лоб (41 поле по Бродман); за соматосензориума
в париеталния лоб (3 поле по Бродман).
От първичните зони на кората сензорната
информация отива към съседни зони –
вторична сензорна кора, а от вторичните
зони информацията се предава към
третичните зони.
Видове гнозис:
1. Зрителен гнозис
– зрителна преработка на информацията.
Аксоните на ганглийните клетки на
ретината образуват nervus
opticus (зрителния нерв).
Първият синаптичен контакт на ганглийните
клетки, образуващи зрителния нерв е в
nucleus geniculatus
lateralis (страничното коленчато
ядро) или в colliculus superior
(горното хълмче). Тъкмо на тези две места
се извършва преработка на зрителна
информация преди да се предаде на
зрителната кора. Има два вида кръстосване
– едно в ретината и едно в хиазмата.
Информацията от дясната половина на
зрителното поле постъпва в лявата
половина на ретината, а от лявата половина
на всяко око – до лявата половина на
мозъка, а чрез кръстосване в хиазмата
– и в дясната половина на мозъка. След
кръстосването (частично) в хиазмата,
аксоните на ганглийните клетки
продължават, като образуват tracticus
opticus. Тук, като се запазва
интакта, информацията се предава от
ретината, без да се преобразува. Tractus
opticus пренася зрителна
информация от половината на всяко око
до 2 структури – colliculus
superior и nucleus
geniculatus. Сolliculus
superior, което е в средния
мозък, имат значение за организацията
на очните ябълки (bulbus
oculi) и за някои перцептивни
функции. Nucleus geniculatus
lateralis е част от таламуса,
където се прекъсват 80% от аксоните на
ганглийните клетки на ретината (другите
20% завършват в colliculus
superior), и предават информацията
на други неврони. Тъй като са след
хиазмата, тези структури получават
информация от двете очи. Екстрастриарната
кора също е включена в сложната преработка
на информацията. Хората с лезия в тези
корови полета на екстратриарната кора
имат нормално зрение и езикова
компетентност, но не могат да четат
думите. Според Томпсън, различните части
от мозъка са включени в преработването
на зрителна информация. При пациенти с
лезии в зрителната кора в окципиталния
лоб има промени в т.нар. периметрия.
Нарушенията, които могат да се регистрират
чрез периметрия са поява на скотоми в
зрителните полета. Когато скотомията
обхваща половината или части от зрителните
полета, се документира хемианопсия
(хомонимна).
2. Слухов гнозис.
Слуховата система е специализирана да
анализира звуците, т.е. вибрациите на
молекулите във въздуха. Човек чува само
тези вибрации, които са между 20 и 2000
херца или трептения в секунда. Възприемането
на гръмкост, височина и тембър на звука
се свързва с амплитуда, честота и
комплексност на трептенията. Звуците
преминават през външното, средното и
вътрешното ухо и активират рецепторите.
Като преминават през слуховите канали,
те достигат до тъпанчето и предават
вибрациите, които се трансформират чрез
3 костици – чукче, наковалня и стреме.
Те се предават на течността в охлюва.
Речевата перцепция е комплекс от процеси,
произтичащи от богатството на речника
на всеки индивид, вариантите на речевите
стимули от техните звукови характеристики
(между говорещи и на говорещия) и
сегментирането на думите една от друга,
фонемата, която е най-малката единица
отличаваща речевия звук от други звуци,
се произвежда на различно място по
различен начин и с различно озвучаване.
3. Тактилен гнозис
– Сетивата са пет зрение, слух, допир,
вкус и мирис. Допирът включва усет за
температура (топло, студено), за докосване
и за равновесен усет. Когато усещаме
допира на предмети, като топка за
пинг-понг, кубче лед или къс хартия, ние
усещаме поотделно техни качества като
температура, форма, размер и пр. Цялото
ни тяло е покрито с кожа, чрез която
можем да възприемаме чрез докосване.
Най-активно използваме ръцете, като
чрез кожата определяме физическите
характеристики на предметите. В кожата
има множество вени, артерии, потни жлези,
косми и рецептори.
Видове АГНОЗИИ:
Зрителната агнозия
е невъзможност да се разпознават видени
обекти; тактилната агнозия –
невъзможността да се опипват обекти;
слухова – невъзможност да се познават
чути обекти (характерните им звуци).
Агнозиите могат да се проявяват в
зрителна, слухова и тактилна модалности.
Вътре във всяка модалност те се делят
и по характера на стимула – за цвят,
форма, физиономия, пространство и пр.
Лазацер описва различни причини за
проява на зрителна агнозия за обекти.
Според него има аперцептивна и асоциативна
агнозия. Аперцептивното нарушение
кореспондира с крайната фаза на чисто
перцептивното ниво на преработка на
информацията.
Терминът аперцептивна
агнозия се използва са случаите на
нарушени елементарни зрителни функции
като острота, различаване, цветово
зрение. Понятието асоциативна агнозия
се определя от 3 критерия. Първо, трудно
се узнават обектите представени по
различен начин: чрез невербални тестове,
чрез назоваване, чрез групиране по
техните семантични категории и чрез
описание на функциите им. Второ, обектите
се разпознават трудно чрез други
модалности освен зрението, напр. чрез
опипване. При всички случаи има перцептивна
абнормност – скотомии, хемианопсия и
пр.
Прозопагнозия –
неспособността на хора да разпознават
лица, включително и своето собствено
лице. Често това е свързано и със
зрителната агнозия. Има три типа
прозопагнозия: 1.нарушение на зрителния
анализ на лицето; 2.нарушение на перцепцията
на лицето; 3.нарушение при разпознаване
на индивидуалното лице.
Има и агнозия свързана
с неспособността да се разпознава
пространството – свързана е с
надмодалностната характеристика на
пространството. В патологията се открива
зрително-пространствена агнозия.
Нарушения се появяват при четене на
отделни думи, при писане, при смятане.
Прерисуване и рисуване обикновено са
невъзможни при пациенти с пространствени
нарушения. Пропускат се или елементи
от пространството, или цялата половина,
разположена в лявата част на пространството.
Измерването на пространствения анализ
може да се извърши с два теста: за
пространствена дискриминация или за
пространствено изследване на по-комплексни
задачи, изискващи памет за пространствена
информация.
Пространствената
дискриминация – използва се тест за
разделяне на линия. Трябва да се определи
центъра на различни по дължина линии.
Пациентът обикновено измества центъра.
Дискриминация на позиция – определя
се позицията в пространството на
двуизмерни прости стимули. Ориентация
– неспособност за определяне на позициите
на две линии като еднакви или различни
в пространството. Номериране – Кимура
изследва пациент, като изисква от него
да назове броя на точки, образи, стимули,
представени бързо чрез тахистоскоп, и
намира значим брой грешки. Пациенти с
церебрални лезии извършват трудно тази
задача.
Гнозис за време – много
древна способност на човека да оценява
времето. Човек се опира на два репера,
между тях ограничава интервал от време.
Времевата перцепция приоритетно се
свързва със слуха. Тъкмо поради това
слуховата модалност се използва
най-интензивно за времева перцепция и
се постулира активното участие на lobus
temporalis.
Гнозисът за ритъм се
нарушава при лезии по-често в дясното
мозъчно полукълбо, понякога и в лявото
и предизвиква амузия (неразпознаване
на музиката). Времевата поредица от
звукови стимули се оценява както чрез
лявата, така и чрез дясната церебрална
хемисфера.
Пространствено игнориране
– използват се тестове с писане,
прерисуване и пр. Установява се едностранно
пространствено игнориране. Наблюдава
се при пациенти с лезия в дясната или
лявата хемисфера.
Праксис
и апраксии
Различните двигателни
нарушения корелират с определена увреда
на мозъка. Например при лезии в първичната
моторна зона се проявяват ограничение
на движенията на крайниците, противоположно
на мозъчната увреда (хемипарализи). При
паркинсонизъм също има нарушение на
двигателната функция макар и по различен
начин. Има обаче друг тип двигателни
нарушения, които са дисфункции на сложни
волеви действия, които не са поради
увреда на първичните моторни зони
(парези) или на съответните сензорни
зони (хипестезии) или на малкия мозък и
др.мозъчни структури за осъществяване
на елементарните движения. Изследването
на нарушенията на волевите действия
включва различни видове задачи –
единични повтарящи се действия; непознати
действия и действия с определена
последователност; познати пози (жестове);
манипулации с предмети и др.
Единични повтарящи
се движения – движения с едната или
с двете ръце. При движенията с едната
ръка се използва тест за докосване от
двете страни на линията, като се регистрира
през определен период от време броят
на докосванията на три етапа. При движения
с двете ръце от изследвания се изисква
да постави ръцете си върху маса, като
извършва симултанни движение с едната
или другата ръка и редува последователно
юмрук или длан – при тези симултанни
движения се наблюдават специфични
нарушения.
Непознати действия
– повторение на единични пози на ръката
(пациентът повтаря единична поза на
ръката – кръг с пръсти, изпънат един
пръст); взаимоотношение между действия
и множество действия (сравняват се групи
пациенти, изпълняващи единични или
множествени последователни действия
– според тях нарушенията се дължат на
паметов, а не на двигателен дефицит).
Познати пози (жестове)
– изследванията доказват, че тези пози
или жестове, които изпълняват известни
комуникативни функции, се разпадат по
различен начин, но не и на манипулациите
с предмети (поздрав, отдаване на чест)
Манипулации с предмети
– манипулации с реални предмети,
пантомимично използване на обектите
(карат пациента да покаже чрез пантомима
използаването на предмет, който му се
показва).
Части на тялото и
специфични действия – получава се
друг вид дефицит – апраксия при ходене,
проявява се и при волеви движения само
на стъпалото или на крака – при ритане
или движение на стъпалото в кръг. При
орална апраксия пациентът не може да
възпроизведе различни орални движения
под команда, независимо че си отваря
устата, когато се храни. Локализация и
латерализация на апраксията на крайниците
– пациентът може да изпълни всеки
определен акт, но не успява да извърши
последователността от актове – при
болестта на Алцхаймер.
Конструктивен праксис
– Това са пациенти, които не могат да
извършват сложни волеви дейности в
пространството, засегнати са всички
движения под зрителен контрол.
Напр.трудност при рисуване на геометрични
форми.
Орална апраксия –
наблюдава се при едностранна хемисферна
увреда, по-често лявата и три пъти
по-малко дясната. Пациентът не успява
под команда да отвори устата си независимо,
че правел това, когато се храни. Описани
са и кинестезичен и кинетичен орален
праксис.
Локализация и
латерализация на апраксията на крайниците
– клас разделяне на апраксиите от
Липман. Идеомоторна, кинетична и
идеаторна. При идеомоторна апраксия
пациентът прави грешки във времето и
пространството. Описани са 3 форми –
при лезия в corpus callosum,
при лезия в предната част на доминантната
церебрална хемисфера и свързана с лезия
в задната част на доминантната хемисфера.
Пациентът в идеаторната апраксия е с
едностранно нарушение на движението.
При нея е засегната ръката.
Конструктивна апраксия
– наблюдава се както при увреда на
лявата хемисфера, така и при увреда на
дясната хемисфера. Тази апраксия се
изучава чрез разнообразни тестове за
рисуване. Лайбман през 1905 определя три
вида апраксия – идеаторна – когато
липсва идея за движение; идеокинетична
(идеомоторна) – при невъзможност за
манипулация с предмети; кинетична
(моторна) апраксия.
Класификация
на афазиите
През 1885 Лихтхайм създава
своята модулна хипотеза за обяснение
на организацията на езика. Според него
има 4 центъра за разпознаване и
възпроизвеждане на езикови структури.
Те са: А – за разбиране на чута (чужда
или своя) реч; В – за възпроизвеждане
на разбрана (правилно възприета) реч; М
– за моторна картина на мускулите, които
осъществяват говор и писане; О – за
зрителна картина на думите (четене и
писане). По-късно се обогатява със
следните елементи. Посочва се съществуването
на мнестично поле (М), което е в двустранна
комуникация със зоната на Брока (В) и
зоната на Вернике (W). Освен
това тези две зони са контролирани от
т.нар. актуално поле (А). Зоната на Брока
се намира в lobus frontalis,
а тази на Вернике – в lobus
temporalis. Въз основа на
модулната хипотеза е изградена
класификацията на Лихтхайм – Вернике
за езиковите нарушения, които могат да
се класифицират като моторни (експресивни)
и сензорни (рецептивни) нарушения.
Моторни (експресивни)
афазии:
| Нарушени | Езикови способности | Съхранени |
| Субкортикална моторна афазия (словесна немота) |
Спонтанна реч
Повторна реч
Четене на глас |
Разбиране на реч
Прочетена
Написана
|
| Кортикална моторна афазия (афазия на Брока) |
Спонтанна реч
Повторна реч
Писане под диктовка и спонтанно
четене на глас |
Разбиране на реч
Чута
Прочетена
|
| Транскортикална моторна афазия |
Спонтанна реч
Спонтанно писане |
Повторна реч
Четене на глас
Преписване, писане
под диктовка
Разбиране на написаното |
Сензорни (рецептивни)
афазии:
| Нарушени | Езикови способности | Съхранени |
| Субкортикална сензорна афазия (словесна слепота, словесна глухота) |
Разбиране на реч:
чута, прочетена
Повтаряне на чути изрази или писане
под диктовка |
Разбиране при писане под диктовка |
| Кортикална сензорна афазия (афазия на Вернике) |
Разбиране на реч:
чута, прочетена
Повтаряне на чута реч, четене на глас |
Спонтанна реч с вербални и литепарафазии, неологизми, „словесна салата” |
| Транскортикална сензорна афазия |
Разбиране на реч:
чута, прочетена
Повтаряне на чута реч, четене на глас |
Писане под диктовка
Преписване без
разбиране
|
Към тези две групи
афазии се прибавя и т.нар. проводникова
афазия. Тя се характеризира с проява на
парафазии. Разбирането е относително
съхранено, както на чута, така и на
прочетена реч.
1973 – Лурия предлава
своя класификация на афазиите, като се
основава на модалностно-специфичната
организация на вторичните асоциативни
полета в церебралните хемисфери. При
лезии в лобус окципиталис в лявата
церебрална хемисфера се проявяват
езикови нарушения, определени като
алексия или латерална агнозия. При лезия
в lobus temporalis
sinistra може да има нарушение
на фонематичния слух (глухота само за
фонеми). Това се нарича акустична афазия.
Ако се прибави и паметов дефицит на думи
– акустико-мнестична афазия.
Гоодглас и Каплан (1972)
правят класификация на афазиите, като
използват анализ на лингвистичните
функции. Те допускат нарушения на
разбирането от слухов и зрителен тип и
няколко експресивни нарушения на
артикулацията, в писането, граматич
структури и потока на езиковата изява.
Има честотен речник за българския език,
има думи подредени по честота на употреба.
Когато тези думи липсват – тежка форма
на аномия. Амнемия (нарушаване на паметта,
забравяне на думите и още амнестична
афазия). Персеверация – ако пациентът
е успял да каже дума или изречение, то
непрекъснато повтаря това. Дефицит на
предните, фронтални глухи зони. Нарушаване
на автоматичната реч – например броене
от 1 до 10, на дните от седмицата. Проводникова
афазия – човек може да говори, но не
може да повтаря чутото и да чете на глас.
Нива
на организиране на езиковата способност
Езикът влияе на
възприятието. Зрителните и слухови
сигнали (разговорна и писмена реч) се
преобразуват от вторичните структури
на мозъка и изпълняват уникална функция
– те не само служат като средство за
кодиране на информацията, но са и част
от филтрираната система, което управлява
количеството и качеството на информацията,
постъпваща за по-нататъшна обработка.
По тези причини различните психологически
клонове изучават езика и неговите
компоненти. Лингвистиката изследва
относително строежа на езика.
Психолингвистика – с психологическия
синтез на 32 основни граматични компоненти
на езика: фонеми, синтаксис, семантика.
Използват теоретичните твърдения в
лингвистиката и когнитивната психология,
изследователите на афазиите изучават
лингвистичните прагове на афазиите и
ги отнасят към субкомпонентите на
системата за езикова преработка. Тези
субкомпоненти са свързани с други нива
на преработка и реализация на езика на
експресивно и рецептивно равнище с
неврологично, праксисно, лингвистично
и идейно ниво.
І. Експресивно равнище
– доминантно съхранен говорен апарат.
Тук участват черепно-мозъчните нерви
и мускули. При патология се получава
псевдобулбарна афазия. Патогенни
рефлекси – болезнено усилване на
рефлексите на главата; устата е леко
отворена и леко се почуква брадата,
автоматично се затварят устните –
централна парализа. Дизартрия – парализа
на говорната мускулатура – първично
двигателно нарушение. Анартрия – не е
езиково лингвистично нарушение.
Праксисно ниво.
Орална апраксия – нарушение на езика
довежда до нарушение на системата за
моторен контрол. Пациентът не може да
възпроизведе орални действия под
команда. Мануална апраксия – нарушение
на движенията за писане.
Лингвистично ниво.
Транскортикална моторна афазия –
лингвистични нарушения, невъзможност
за използване на лингвистичните правила.
Когато пациентът слуша и чете голям
приток от езикова информация и не разбира
смисъла й, говорим за транскортикална
сензорна афазия – няма гнозисни
нарушения. Следващото нарушение е афазия
на Брока на праксисно и лингвистично
ниво.
Идейно ниво.
Говорейки, има идея какво да каже, но не
може да подреди правилно изречението
(не е афазия, а друг вид езиково нарушение.
Психоза.)
ІІ. Рецептивно равнище
– за разбиране и възприятие на езикова
информация чрез зрителна, слухова и
тактилна модалност
Неврологично ниво
– езиковото построяване е нормално, но
нарушение има. Слепият чете опипвайки,
глухият говори жестикулирайки;
хематоанопсия.
А) Слухова модалност –
фонематичен гнозис. Патология – при
нарушения в мозъчната кора (афазия),
невъзможност да се разбере чутата реч,
не се компенсира с други сетива;
Б) Зрителна модалност
– буквен гнозис (виждане на езиковите
знаци). Патология – има дефицит за букви
– става трудно разбираема написаната
информация, но чутата реч е разбираема.
Праксисно ниво –
не се разбират лингвистичните правила
и пациентът не знае как да се произнесе.
Познанието на езика ни дава възможност
да разберем, че патологичните причини
при езиковите способности се базират
на недоразвитието на вторичните и
третичните функционални зони.
Идейно ниво –
психоза. Своят интерпретация на зрително,
слухово и тактилно равнище, без да имаме
езиково нарушение въобще, а само дефицит.
Лингвистично ниво
– транскортикална сензорна афазия на
Вернике – на гнозисно и лингвистично
ниво. Формално е съхранено по-елементарното
езиково разбиране, но с пациента трудно
може да се осъществи контакт (нито
парези, нито парализи).
Калкулия
и акалкулия
Способността за
изчисляване съществува в човешката
популация, независимо от обучението в
училищата – събиране, изваждане, смятане
на пари, на календар и др. Способността
да се изпълняват рутинни, аритметични
изчисления се нарушава селективно при
пациенти с церебрални лезии. Дефицитът
при калкулацията е свързан и с други
езикови трудности. Според Неншен
специфичното нарушение на калкулацията
трябва да се нарича акалкулация. Бергер
открива 2 вида дефицит – първична и
вторична акалкулация. Първичната е
акалкулация, при която нарушението е
изолирано, а вторична е, когато е
придружена с нарушения в четенето, езика
и др. Определени са следните видове
акалкулия – анаритметрия – първичен
дефицин на калкулацията, съответстващ
на класификацията на Бергер; алексична
и/или аграфична акалкулия – когато има
специфичен дефицит при четене и писане
на числата; пространствена акалкулия
– когато се появяват грешки поради
пространствена дезорганизация на
писмената калкулация.
Разбиране и
възпроизвеждане на цифрите – разбрани
и проговорени цифри – тежко нарушение
при проговоряне на разпознатите цифри,
без други дефицити. Разбрани и написани
цифри – например не могат да се разпознават
5 и 7, а също и думите, означаващи 5 и 7.
Това е селективно проявен дефицит.
Дисоциация може да има в разбирането
на написаните цифри – нарушение за 5 и
7, а не за „пет” и „седем”, и обратно.
Нарушение може да има и при разпознаване
на знаци (*,/,+,-,=). Извличане на цифри и
числа – пациентът прочита правилно
цифрите написани със знак или дума
(например 4+5 или „четири плюс пет”),
произнася „8”, написва „5” и избира
„9” за верен отговор. Алексия за числа
– използва се тест за четене на арабски
цифри. Пациентите прочитат отделните
цифри, но не и числото съставено от тях.
Разбирането и извличането
на цифри се извършва чрез левия париетален
лоб, от части от g.angularis.
Четенето и писането също се нарушават
при лезии в тези зони. Голяма роля за
калкулацията има лявата церебрална
хемисфера. Според Бентън увреда в левия
париетален лоб предизвиква акалкулия.
Калкулацията е сложен
мултикомпонентен процес. Необходимо е
първо разпознаване на цифрите за
калкулация, актуализиране на действието
и извличане на конкретната дума – цифра
(число) за отговор. Процесът думи-цифри
е подобен на този четене-букви. Намерени
са зрително-пространствени нарушения,
които увреждат писането на ифри, но това
се отличава от първичния дефицит за
калкулация. Първична калкулация се
проявява при интактна способност за
пространствен анализ. Анатомичния
дефицит при пространствени нарушения
е свързан с дясно хемисферна дисфункция,
а изпълнението на тестовете за аритметично
проговаряне – с ляво хемисферна лезия.
При първична акалкулия има нарушения
на знанието за аритметичните процедури
и нарушение на знанието за аритметични
факти.
Памет
и амнезии
Краткосрочна памет
– в паметта си вербален материал можем
да удържим толкова дълго, колкото е
необходимо до точното му възпроизвеждане.
Когато се изучава краткосрочната памет
е прието да се изследват границите на
капацитета и на продължителността за
запомняне на инфомация. Изучаването на
капацитета включва броя айтеми или ниво
информация, която се удържа и възпроизвежда
след единично представяне. Изследването
на продължителността на краткосрочната
памет фокусира вниманието, за което
материалът е забравен, ако липсва влияние
на вниманието.
Слухова КП –
модалностно-специфична и материално-специфична.
Капацитет – за здрави индивиди капацитетът
на краткосрочната памет не се определя
от физическата същност на стимулите.
Продължителност – КП не е свързана само
със съдържанието на информацията, но и
с времето за нейното задържане. Например,
ако искаме да си припомним непознат
адрес или телефон, си ги повтаряме наум,
с идеята да ги запомним или да задържим
най-дълго време информацията за тях.
Материално-специфични ефекти – трудно
запомняне (удържане на слухово-вербален
материал) при пациенти се дължи на
нарушение, свързано с обема и с интервала.
При запомняне на един невербален слухов
материал също се наблюдават проблеми.
Зрителна КП – при
наблюдавано селективно нарушение за
запомняне на слухов материал, запомнянето
зрително е съхранено. Наблюдавани са и
обратните селективни нарушения – за
зрително представен материал е нарушено
запомнянето, а за слуховата е запазено
(Зрително-вербален материал се
експонира с кратка продължителност на
стимулите, в порядъка 100-200 м/сек. Видяното
за толкова кратко време се възпроизвежда
чрез назоваване. При нормални хора се
възпроизвежда за 4-5 айтема.
Зрително-пространствен материал –
използват се при пространствено-организирани
стимули за кратко показване на пациенти.
Айтемите са едни и същи, но варира тяхното
разположение в пространстовот. Нормално
се запомнят 5-6 позиции от тях.)
Автобиографична и
дългосрочна памет – с времето много
от спомените нормално се модифицират,
разнообразяват или се забравят.
Възпроизвеждането на събития случили
се вчера са с повече детайли от тези
преди седмица или от други случили се
в минали години. Възстановяването на
автобиографич събития е възможно
благодарение на дългосрочната памет.
Нарушаването на способността за
използването и възпроизвеждането на
адекватна автобиогр информация е в
зависимост от церебралното заболяване.
В т.нар. ДП нарушенията се разглеждат
като глобално увреждане на АП или
нарушение, засягащо способността на
пациента да си припомня известни групи
от случки през годините (материално-специфични
нарушения). Глобални нарушения на АП –
пациентът има дефицит за минали срещи
и бъдещи събития. Субтипове на загуба
на паметта (amnesia) – чист
синдром на амнезия е забравяне на всичко,
отнасящо се до него. В своята чиста форма
амнезията е проучвана интензивно през
годините. В края на ХІХ в. Корсаков
отбелязва два компонента на синдрома
амнезия. Първо, пациентът трудно
възпроизвежда и узнава събития, които
са се случили след заболяването –
антероградна амнезия; второ, пациентът
има нарушено възпроизвеждане на събития,
случили се преди причината за амнезията
– ретроградна амнезия.
Формула на трите R
1. Регистрация –
когато трябва да се проучва паметта,
трябва да се каже какъв вид материал
трябва да се даде на изследваното лице.
Когато искаме да изследваме памет с
невербални стимули, се използва формулата
на Милер – 77 1+2 айтема. Материалът, който
се подава за регистрация е 5,6,7,8,9 айтема.
Това се получава за невербални и вербални
стимули. Когато тази формула е 5, то
тогава е патологичен случай. Трябва да
се уточни чрез коя сензорна модалност
се изследва паметта (уточнени паметови
айтеми) и да се уточнява за коя модалност
става въпрос. Паметта е еднакво развита,
независимо за коя сензорна модалност.
2. Ретенционен интервал
(задържане на информацията) – сензорна
зрителна памет, слухова сензорна памет
– ехонична, кинезистатична сензорна
памет, КП – от 20-30 сек, ДП.
3. Retrieval
– проверка на паметта. Тази проверка
се прави чрез два варианта – а)узнаване
– подават се три думи, след това още 3,
още 3. Между първата и третата се връща
във втората, за да се познае дадената
дума; б)възпроизвеждане – свободно
възпроизвеждане, серийно възпроизвеждане
по реда, по който е дадена информацията.
Ако това е на много ниско равнище, този
вид проверка дава много ниски стойности
на паметта.
Видове амнезии:
1. Антероградна амнезия
– пациенти с антероградна амнезия не
са само с нарушено възпроизвеждане
(recall), но трудно изпълняват
и задачи за узнаване. След тестове се
установява глобален дефицит за вербална
и невербална памет. Пациентите изпълняват
твърде лошо общоприети тестове за памет.
Двигателните умения се използват при
обучение на пациенти с амнезия, например
рисуване под контрол; използва се и
перцептивно обучение, например да се
прочете и научи някакъв текст. Използва
се запомняне на думи (чрез списък с
думи); вербално усвояване. Пациенти с
амнезия могат да имат проблеми и с КП –
това се случва при задачи за незабавно
възпроизвеждане.
2. Ретроградна амнезия
– невъзможност за възпроизвеждане на
събития, които са се случили преди
появата на самата амнезия. Пациенти с
органична амнезия имат слабо възпроизвеждане
за много събития от живота си. АП се
изучава чрез формално изследване за 4
периода от живота – училище, работа,
женитба, семейство. Тези задачи се
повтарят през 7 дни. Изучаването на
ретрогр амнезия става чрез възпроизвеждане
на по-стари спомени. Парамнестични
нарушения – нарушения, при които
пациентът може да създава „спомени”,
които не са свързани с „актуални”
събития от миналото. Конфубулациите
могат да бъдат фантазии и предизвикани.
При пациенти с церебрални лезии се
отличават 2 вида нарушения на паметта
– на вербалната памет (за възпроизвеждане
на имената на хора, хотели и пр.) и на
невербалната памет (за местонахождание
на улици, площади и пр.). И двата вида
памет се изучават чрез различни тестове,
напр. на Милер, Бентън, Кимура. Има още
памет за физиономии и топографска памет.
Има предположение за
два различни пътя за невроналните основи
на амнезията. Единият е, че всяка амнезия
има количествени промени, разликите са
само в степента на проява на дефекта
или са в резултат на увреда на част от
мозъка, която не е директно включена в
амнезията. Другият път е, че увредата в
различни части на паметовата система
води до количествени разлики във формата
на амнезията. Явяват се нарушения на
медиалната повърхност на римооралния
лоб и на медиалната част от диенцефалонната
зона, т.е. два вида амнезия според
локализацията на лезиите. Резултати от
две групи експерименти доказват, че
медиотемпоралната група е с акселерация
на забравянето, а медиодиенцефалната
– няма такава. Хипокампът и свързаните
с него структури са специализирани за
проучване, съхраняване и извличане на
информация.
Функционална
мозъчна асиметрия
Двете мозъчни хемисфери
не са симетрично огледални образи една
на друга. Тяхната асиметрия започва от
ембрионалното развитие и при раждането
в около 65% от индивидите, повърхността
на слепоочния коров дял на лявата
хемисфера е по-голям. Тази вродена
структурна асиметрия предопределя и
по-благоприятното следродово развитие
на лявата хемисфера – към нея постъпва
по-богат поток от стимули, което от своя
страна благориятства последващото
по-силно развитие на тази мозъчна част.
Във връзка с това 90% от индивидите се
оформят с по-голяма сръчност на дясната
ръка. Най-подчертана е мозъчната асиметрия
по отношение на езиковите функции.
Езиковите зони в преобладаващия процент
от хората са разположени само в лявата
хемисфера и това я определя като
доминантна или категорийна хемисфера.
Дясноръкост и леворъкост не се припокрива
с латерализацията на категорийната
хемисфера. При 99% от десноръките лявата
хемисфера е категорийна. При леворъките
дясната хемисфера е категорийна само
при 30-40% от случаите. При леворъките
езиковите функции са ясно латерализирани,
поради което смущения в говора се
появяват при увреждане на която и да е
страна на мозъка. Историята на локализацията
на езика и на латерализацията на функциите
започва с Брока, когато долната част на
лявата префронтална мозъчна кора (зона
на Брока) се определя като център на
речевата продукция. Това се дължи на
gyrus precentralis,
контролираща мускулите на устната
кухина и лицето. Увреда в зоната на Брока
води до разпадане на речевата продукция,
без да има дефицит в езиковото разбиране.
Вернике посочва втори кортикален езиков
център – зона на Вернике – лява темпорална
кора, точно задната част на първичната
слухова кора, и е за разбирането на
речта. Според него лезия в зоната на
Брока предизвиква афазия на Брока
(моторна афазия). Лезия в зоната на
Вернике води до сензорна афазия.
В първите години на
ХХв. Липман открива апраксията. Тя се
свързва с лезия в лявата церебрална
хемисфера. Пациентите имат трудности
да изпълняват действия по програма,
т.е. имат двигателни или сензорни
дефицити, а симптомите се проявяват в
билатерално-доминантна хемисфера.
Ако се приеме, че лявата
хемисфера доминира за всички значими
дейности, то се открива, че дясната
хемисфера е приоритетна за много задачи,
изискващи ориентация в пространството,
изискващи ориентация в пространството.
Лявата ръка и дясната хемисфера са
по-бързи и по-точни, докато дясната ръка
и лявата хемисфера често се придружават
от вербален коментар, потискащ трудностите
на изследваното лице. Дясната хемисфера
е с приоритет за емоции, както за
емоционална реакция на различни стимули
и ситуации. Има сензомоторна теория на
мозъчната асиметрия, на Кимура – според
нея лявата хемисфера не е специализирана
за контрол на речта, а на фините моторни
действия, от които речта е само една
част. Лезии в лявата церебрална хемисфера
често нарушават волевите устни действия,
което не съпътства речта. При лявохемисферна
лезия се проявява афазия. Аналитико-синтетичната
теория за церебралната латерализаця
идва като неочакван факт. Тя се опитва
да изясни защо речта е латерализирана
към лявата хемисфера. Това е, защото
сензомотроното умение е латерализирано.
Уишам и Колб описват
разлики в невроналната структура на
лявата и дясна церебрални хемисфери.
Първото съобщение за невроанатомична
асиметрия е от 1800, напълно игнорирано
до съобщението на Левитски, 1968, за
по-обширен planum temporale
в ляво в сравнение с този в дясно при
65% от човешките мозъци. Според Вада и
сътрудници, 1975, изследвали мозъка на
фетуси, planum temporale
е по-широк. Те предполагат, че левият
темпорален лоб е структура за специализация
на езиковата активност още преди да се
развие речта. Зоната на Брока в лявата
хемисфера също е по-обширна вляво и
вдясно не винаги корелират с езикова
функция. Напр. 95% от хората са с
лявохемисферна доминантност за речта,
а само при 65% planum temporale
е по-обширен вляво.
Методите за изследване
на функционалната мозъчна асиметрия
са инвазивни и анвазивни.
Методи
за изследване на функционалната мозъчна
асиметрия – инвазии и анвазии
Проба на Wada
(Sodimn Amytal Test) – това е тест за
езикова латерализаця, инвазивен и
провеждан само при пациенти по медицински
индикации. В неврохирургията се използва
проба на Wada за определяне
на езиковата хемисфера на всеки пациент,
на който му предстои операция в нея.
Инжектира се контрастно вещество във
вътрешната сънна артерия, за ад се
направи ангеография на мозъчни съдове.
След това се включва наркотизиращо
вещество. За няколко минути се анестезира
тази половина на мозъка, от която е
инжектирана вътрешната сънна артерия,
докато капацитетът на другата хемисфера
е нормален. По време на анестезията от
пациента се изисква да повтори добре
позната информация (напр. азбуката),
както и да назове познати обекти. При
различни пациенти се проучват възможностите
или на лявото или на дясното полукълбо.
Ако инвазираната част е доминантна за
речта, пациентът има редица дефицити
при изпълнение на горепосочените задачи.
При обратния случай – хемисферата не
е доминираща – в речта има малко грешки.
Чрез този тест Милнер открива, че 92% от
десноръките са с лявохемисферна доминанта
на речта, докато 69% от леворъките и
амбидекстрите имат доминанта за речта
лявата хемисфера. При 30% от тях лявата
хемисфера не е доминантна.
Тест за дихотично
слушане (Dichotic
Listening Test)
– проучванията на пециенти с едностранна
мозъчна увреда и чрез пробата на Wada
дават сведения за речевата латерализация.
Но според Кимура, изследванията на
здрави хора изискват специална методична
постановка. Използва се тест за дихотично
слушане, който се състои от 3 двойки
едновременно изговорени чрез магнетофон
думи, като всяко ухо слуша един от двата
стимула. Напр. от едната страна се чува
3,5,7, а от другата 1,6,8. Изизсква се да се
назове всяко чуто число. Открива се, че
най-много хора назовават числата, чути
с дясното ухо, а доста по-малко са тези,
които назовават чутото с лявото ухо. От
тух се прави извода, че доста хора имат
доминанти за езика – лява церебрална
хемисфера. Тя приеротетно реагира на
информация, приета с дясното ухо. За
доказателство се изследват и пациенти,
за които е доказано чрез проба на Wada,
че имат дяснохемисферна доминанта. Те
изпълняват теста за дихотично слушане
с приоритет – ляво ухо.
Други тестове за
езикова латерализация при здрави хора
– след теста за дихотично слушане се
създават и други тестове за проучване
латерализацията на здрави хора. Напр.
за зрителната модалност се използва
тахитоскопична техника. Изследването
се основава на принципа да се сравнява
възможността за идентификация на думи,
представени в лявото или в дясното
зрително поле. Изследваното лице фиксира
с поглед центъра на екрана, а всяка дума
симултанно се експонира за 0,1 сек. Времето
за узнаване на думата е по-кратко от
инициирането на очните движения. За
четящите в посока от ляво на дясно,
думите се изследват при вертикално или
хоризонтално представяне с различни
конфигурации на графемите (заострени,
заоблени) и в сравнение с безсмислените
съчетания от букви. По-бързо се разпознават
и назовават думите, видени чрез дясното
зрително поле – лявата церебрална
хемисфера. Така е при тези хора, които
са с доминантност на лявата церебрална
хемисфера за езика (повечето от
десноръките). При идентификация на
неезиковите стимули не се доказва
приоритет на дясното зрително поле, а
на лявото – т.е. стимулите се възприемат
приоритетно от дясната церебрална
хемисфера.
За доказателство на
езикова латерализация се изследват и
движения, съпътстващи речта. Тай като
фините движения се контролират от
контралатералната хемисфера, Кимура
намира активност на дясната ръка при
доминантност за езика на лявата хемисфера,
е специализирана за контрол на комплексни
двигателни дейности на ръката и на
устните при речева дейност.
Изследвания са правени
и за дихаптично представяне на вербални
стимули за опипване едновременно с
двете ръце (активен или пасивен допир).
Подобно на дихотичното слушане и на
тахистоскопичното латерализирано
изследване и чрез дихаптичната моторика
се доказва, че приоритетно се разпознават
езикови стимули опипани чрез дясната,
а разпознаването на безсмислени стимули
е по-добро чрез лявата ръка.
И по трите методики
(дихотична, дихаптична и тахистоскопична)
данните са различни (за езикова
латерализация и неезикова преработка).
Зависят и от други характеристики на
представения стимулен материал или от
промени в процедурата на изследването.
Чрез ПЕТ (пезиторна емисионна томография)
се извършва почти революционна промяна
в изучаването на церебралната локализация
на езика. Като използват ПЕТ, Петерсън
и сътрудници, 1990, измерват кръвотока на
различни части на мозъка по време на
езиково-свързани умения. Тестовете по
време на ПЕТ са 10-минутни. Инжектирането
на радиоактивното вещество е интравенозно,
а не в сънната артерия – двете хемисфери
могат да се изучават симултанно. Използва
се компютърна програма за промени в
кръвотока по време на една или друга
активност. Измерванията са правени при
две условия – зрително и слухово подаване
на прогресивно усложняване на задачите.
Електрошокова терапия
– прави се инвазивно, едностранно, за
да се проучи функционалната мозъчна
асиметрия. Инвазивните технологии не
се правят заради експеримент и
психологическо изследване, а за лечение
напр. на шизофрении, епилепсия. Анвазивните
методи (тестове) се използват при здрави
хора.